
Le plus grand ensemble de tables dans FishBase est en rapport avec l'écologie trophique des poissons et présente des informations sur l'habitat, les aliments, le régime alimentaire, la consommation de nourriture et les prédateurs. La table ECOLOGY de Froese et al. (1992) couvrait aussi les limites de tolérance environnementales et des informations sur le comportement, mais bien peu d'ensembles de données convenablement standardisées avaient été trouvés et ces informations saisies à l'origine sont maintenant contenues dans le champ Remarks de la table SPECIES.
Les informations sur l'écologie trophique qui peuvent être utilisées pour la construction de modèles Ecopath (voir Encadré 19), sont présentées dans les tables suivantes :



De plus, une structure hiérarchique a été implémentée pour ces tables pour ordonner les aliments en niveau croissant de détails, du champ Foods I (consistant en 6 types généraux de nourriture différents, listés dans un champ à choix multiples) au champ Food III (55 types de nourriture), avec un champ intermédiaire Food II (22 types de nourriture). Cette structure qui distingue des niveaux (pour les plantes et les animaux), permet aussi l'entrée des informations sur les aliments (par exemple le nom de l'espèce ingérée). Plus de détails sont fournis ci-dessous sur cette structure, ainsi que sur les niveaux trophiques assignés aux différents aliments dans les champs Food I, II et III (voir ‹ La table FOOD ITEMS ›, ce volume) qui peuvent servir à estimer le niveau trophique des poissons dont le régime alimentaire est connu.
| Encadré 19. La modélisation par le modèle
Ecopath et FishBase. Développé dans les
premières années 1980 par J.J. Polovina et ses
collègues au Laboratoire du NMFS à Honolulu, et
appliqué tout d'abord à un système de récif corallien
au nord d'Hawaï (Polovina 1984), l'approche Ecopath pour
la réalisation et l'analyse des modèles de relations
trophiques équilibrées dans les écosystèmes a ensuite
été développée par d'autres auteurs. En particulier,
nous l'avons étendu pour inclure toute une gamme de
routines analytiques, et encouragé son application à
divers écosystèmes (Christensen et Pauly 1992,
1993 ; Pauly et Christensen 1993 ; Christensen
et Pauly 1995 ; Pauly 1997). L'approche Ecopath se
décompose dans les étapes suivantes : 2. Pour chaque groupe fonctionnel i, obtenir
des estimations préliminaires pour tous les paramètres
sauf un de l'équation principale d'Ecopath : Bi
· (P/B)i · EEi = Yi + 3. Utiliser les diverses routines d'Ecopath pour résoudre le système d'équations linéaires dans (2) pour l'écosystème complet. 4. Utiliser le réseau de flux défini par ce système d'équations pour estimer des paramètres tels que des efficacités de rendement par niveau trophique (voir Encadré 20), des indices de sélection de niche, des estimations de la mortalité naturelle (voir ‹ La table POPGROWTH ›, ce volume), etc. Ecopath et FishBase ont plusieurs caractéristiques
communes, en particulier leur objectif de relier
l'halieutique aux disciplines proches ; leur large
disponibilité ; un vaste réseau d'utilisateurs et
de collaborateurs ; et par eux, la mise à
disposition de standards pour leur discipline
respective : la modélisation d'écosystèmes, dans
le cas d'Ecopath : l'ichtyologie appliquée dans le
cas de FishBase. |
Nous sommes conscients que ces tables décrites ci-dessous avec plus de détails ainsi que la hiérarchie des aliments que nous avons créées peuvent paraître arbitraires à première vue. Cependant, comme pour le reste de la structure de FishBase, elles se sont dégagées au fil des améliorations successives résultant des tentatives pour intégrer des données extraites d'une grande variété d'études. Nous pensons qu'elles sont raisonnablement stables dans leur forme présente pour intégrer les nombreuses données que nous avons prévu de saisir.
Remerciements
Nous remercions vivement Pascualita Sa-a pour ses suggestions d'amélioration à la table DIET, et à R. Froese pour son intérêt pour l'écologie trophique et pour son soutien à son intégration dans FishBase.
Référence
Froese, R., M.L.D. Palomares et D. Pauly. 1992. Draft user's manual of FishBase, a biological database on fish. (ver. 1,0). ICLARM Software 7, pag. var.
Maria Lourdes D. Palomares et Daniel Pauly
La première section de la vue sur cette table présente simplement les habitats et consiste en une suite de champs ‹ oui/non › qui indiquent principalement les préférences environnementales de l'espèce. Cette standardisation catégorielle stricte permet de condenser des descriptions textuelles bien trop longues et de construire des requêtes rigoureuses.
Les habitats sont répartis ici selon les types majeurs de salinité, et indiqués sous forme de champs ‹ oui/non ›. Pour les eaux douces sont listés : rivières (Streams), lacs (Lakes) et grottes (Caves), et dans ce dernier cas, un quatrième champ sélectionné ‹ oui › seulement si l'espèce vit exclusivement (exclusively) dans des eaux souterraines. Pour les eaux saumâtres, un champ groupe Estuaires/lagunes/mers saumâtres (Estuaries/lagoons/brackish seas) et inclut les embouchures des rivières. Pour les eaux salées, les champs sont groupés en zone de balancement des Marées (Intertidal) et zone Marine (Marine). Deux types de substrats sont détaillés pour les zones à marées, les fonds meubles (soft) (sablonneux, boueux, vaseux), et les fonds rocheux (rocky).
Pour les zones franchement marines, des catégories ont été établies en fonction de l'éloignement à la côte [province océanique (oceanic), province néritique (neritic)], du type de substrat [fonds meubles (soft), fonds durs (hard bottom)], et du type de colonisation du substrat [récifs coralliens (coral reefs), herbiers ou prairies sous-marines à phanérogames (sea grass bed), forêts sous-marines de macro algues (macrophytes)]. Un champ mangroves/marais/marécages (mangroves/marshes/swamps) regroupe ces habitats qui peuvent être baignés par des eaux de différentes salinités. De plus la zone climato-géographique préférentielle de l'espèce est indiquée parmi les choix suivants : tropical, subtropical, tempéré, boréal, austral, profondeur, altitude.
Nous ne sommes pas très satisfaits de ces classifications qui paraissent pourtant simples, mais qui ne sont pas encore suffisamment opérationnelles, au point d'empêcher de faire des choix clairs pour de nombreuses espèces. Nous apprécierions des suggestions pour des approches plus simples, et cependant plus rigoureuses pour classer les habitats aquatiques.
La section suivante présente des informations générales sur les habitudes alimentaires de l'espèce. Le champ nourriture principale (Main food) indique le type de nourriture principalement consommé parmi les six aliments suivants : détritus ; plantes ; zoobenthos ; zooplancton ; necton ; autre. Le choix ‹ autre › est sélectionné quand le type de nourriture principale ne se trouve pas parmi les choix disponibles, l'aliment spécifique étant alors précisé dans le champ remarques (Remarks).
Le type d'alimentation (Feeding Type) donne une idée générale du niveau trophique occupé par l'espèce dans un réseau trophique (voir aussi Encadré 20) parmi les choix suivants : principalement carnivore, omnivore, principalement herbivore. Ainsi, un consommateur primaire est-il principalement ‹ herbivore › un consommateur secondaire, tertiaire, etc. principalement ‹ carnivore › ; les poissons qui sont à la fois carnivore et herbivore sont enregistrés comme ‹ omnivore ›.
| Encadré 20. Le régime herbivore, un phénomène
des basses latitudes. La table ECOLOGY utilise un
champ à choix multiple pour indiquer grossièrement la
niche trophique des poissons, et
‹ principalement herbivore › est un
des choix. De même, une valeur proche de 2 dans le champ
troph (troph - 2 s.e. La figure 29 tire partie de cette information et présente un histogramme des pourcentages de poissons herbivores par intervalles de 10º de latitude. Elle illustre le fait que les espèces herbivores ont tendance à être plus fréquentes aux basses latitudes, tendance bien réelle malgré leur très faible pourcentage total parmi tous les poissons (>1,1%). Ces deux phénomènes, le faible nombre de poissons herbivores et leur préférence pour les eaux tropicales, peuvent s'expliquer par les difficultés que la plupart des poissons ont à établir et à maintenir, pendant et après leur alimentation, les faibles valeurs de pH requises pour la digestion des matières végétales, surtout à basse température. Le symbole > avant le 1,1 % fait référence au fait que le type d'alimentation n'a pas été complété pour toutes les espèces, et que la valeur par défaut est principalement carnivore. Mais nous prévoyons que le pourcentage d'herbivores ne dépassera pas 2% même quand le champ sera complété pour toutes les espèces, et que le pic aux basses latitudes restera inchangé. Daniel Pauly |

Fig. 29. Pourcentage d'espèces herbivores par latitude chez les Cichlidae et chez les autres poissons. Voir Encadré 20 pour une interprétation de ce graphique.
Un autre attribut important pour les poissons disponible dans la table ECOLOGY est leur niveau trophique (ici abrégé ‹ troph ›) qui définit leur position dans un réseau trophique (voir Encadré 21). Les valeurs de troph peuvent être estimées par plusieurs méthodes. La table ECOLOGY a ainsi deux groupes de deux champs contenant des valeurs de troph et leur erreur-standard (s.e.) : un groupe pour des valeurs issues d'Ecopath (voir Encadré 19) ; un autre extrait de la table du régime alimentaire. Dans les deux cas, la valeur est soit la seule disponible, soit la médiane de plusieurs valeurs. Les estimations de troph dans la table ECOLOGY s'appliquent aux juvéniles/adultes ou aux adultes sauf indication contraire. Un graphique (Fig. 30) peut être affiché pour illustrer la relation du troph médian en fonction de la longueur maximale chez les poissons.
| Encadré 21. Les niveaux trophiques des
poissons. Les niveaux trophiques (ici abrégé ‹ troph ›), correspondent à la position des poissons ou des autres organismes dans le réseau trophique. À la différence des nombres de rayons des nageoires,
les trophs ne sont pas des descripteurs des organismes
eux-mêmes, établis d'après une classification des
types d'alimentation, mais des attributs de leurs interactions
avec les autres organismes. Aussi, pour estimer les
valeurs de troph des poissons, devons-nous considérer à
la fois leur régime alimentaire, et les trophs des
aliments. Le troph d'un groupe donné de poissons
(individus, population, espèce) est alors estimé de la
façon suivante : |
L'autre information importante de cette section est l'habitude alimentaire. Les choix disponibles sont ordonnés selon les positions que les poissons occupent dans la colonne d'eau. La plupart des espèces pélagiques sont soit des prédateurs ‹ prédateurs de necton › dans la colonne d'eau, soit des ‹ filtreurs de plancton › en surface, soit des ‹ brouteurs sélectifs de plancton ›.

Fig. 30. Relation entre le niveau trophique et la longueur maximale chez les poissons. Noter la pente positive indiquant que les grandes espèces ont tendance à être plus piscivores que les petites.
Comment y arriver
Cliquer sur le bouton Ecology dans la vue SPECIES. Le graphique de la figure 25 [troph = f(Lmax)] est accessible de la vue ECOLOGY ou de la fenêtre GRAPHS.
La table FOOD ITEMS
La table FOOD ITEMS contient les aliments dont la consommation a été documentée pour les espèces de poissons. Dans la vue correspondante, ces aliments sont noircis au sein d'une liste prédéfinie pour l'espèce sélectionnée (voir ci-dessous). Cliquer sur un bouton noirci dans le champ Aliment II (Food II) ouvre la vue FOOD ITEM DETAILS qui affiche un ensemble d'informations sur l'aliment, sur le Groupe, le Nom, la Phase du cycle de vie ou la partie du corps ingéré, la Fréquence dans le régime alimentaire, le Pays d'origine de l'échantillon et des Remarques.
| Encadré 22. La hiérarchie des aliments. Pour standardiser les champs des tables d'écologie trophique, une hiérarchie des aliments a été établie du champ Aliment I (Food I : 6 choix, en gras), puis Aliment II (Food II : 22 choix en romain) jusqu'au champ Aliment III (Food III : 55 choix, en italique) :
La table FOOD ITEMS peut être consultée pour les niveaux trophiques assignés à ces divers groupes. Maria Lourdes D. Palomares, Pascualita Sa-a et Daniel Pauly |
Les informations contenues dans la table FOOD ITEMS sont utiles pour définir les relations prédateur-proie des poissons. La compilation des différents aliments consommés par espèce peut permettre la mise en évidence de préférences alimentaires stables dans les régimes des espèces pour lesquelles des données détaillées manquent, et contribuent aux évaluations préliminaires de niveaux trophiques (voir Encadré 21).
Sources
Plus de 400 références ont été utilisées dans la table FOOD ITEMS. Parmi elles, Hiatt et Strasburg (1960), Randall (1967), Scott et Crossman (1973), Allen (1985), Randall (1985), Whitehead (1985), Hickley et Bailey (1987), Maitland et Campbell (1992) et Sierra et al. (1994).
La vérification de plus de 6 600 enregistrements dans cette table, qui concernent plus de 1 500 espèces, a été faite en contrôlant les affinités taxinomiques des aliments. Quelques groupes d'animaux utilisés comme nourriture occupent plusieurs habitats. Des résultats contradictoires dans la classification fonctionnelle de certains aliments en ont découlé. Par exemple, sans autre précision, ‹ copépodes cyclopoïdes › peut désigner des espèces benthiques ou planctoniques. Dans ces cas, nous avons déduit le groupe fonctionnel d'un aliment d'après l'habitat et le comportement de l'espèce qui le consomme.
Les champs
Pour standardiser les données dans la table FOOD ITEMS et les autres tables d'écologie trophique, une structure hiérarchique a été implémentée avec trois niveaux de précision (Aliment I, II, III -Food I, II, III) pour les enregistrements (Encadré 21). Double-cliquer dans la vue FOOD ITEMS sur un bouton du premier niveau Aliment I (Food I), même non sélectionné, ouvre la vue FOOD TROPHS. Pour chaque niveau, elle affiche les valeurs estimées des Trophs (+/- 1 e.s.) qui permettront de calculer ceux pour les poissons dont le régime alimentaire est connu (voir ‹ La table DIET ›, ce volume). Les sources des valeurs estimées du troph sont indiquées dans Référence (Reference). Remarques (Remarks) contient des informations supplémentaires. Il indique notamment si le troph est calculé en utilisant des trophs d'autres groupes, auquel cas, aucune référence de source n'est précisée.
Dans la vue FOOD ITEMS, les boutons sélectionnés de Aliment I (Food I) et noircis de Aliment II (Food II) indiquent les groupes d'aliments qui sont consommés. Cliquer sur un des boutons noircis Aliment II (Food II) affiche une liste des aliments classés par catégories. Double-cliquer sur l'un d'eux ouvre la vue FOOD ITEM DETAILS qui affiche des informations sur cet aliment dans les champs suivants qui indiquent :
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Trophic Ecology dans la vue BIOLOGY, et sur le bouton Food Items dans la vue TROPHIC ECOLOGY. Double-cliquer sur les boutons Aliment I (Food I) dans la vue FOOD ITEMS pour obtenir la vue FOOD TROPHS. Double-cliquer sur les boutons Aliment II (Food II) dans la vue FOOD ITEMS pour obtenir la vue FOOD ITEM DETAILS.
Références
Allen, G.R. 1985. FAO species catalogue. Vol. 6. Snappers of the world. An annotated and illustrated catalogue of lutjanid species known to date. FAO Fish. Synop. 6(125) : 208 p.
Hiatt, R.W. et D.W. Strasburg. 1960. Ecological relationships of the fish fauna on coral reefs of the Marshall Islands. Ecol. Monogr. 30(1) : 65-126.
Hickley, D. et R.G. Bailey. 1987. Food and feeding relationships of fish in the Sudd swamps (River Nile, Southern Sudan). J. Fish Biol. 30 : 147-159.
Maitland, P.S. et R.N. Campbell. 1992. Freshwater fishes of the British Isles. Harper Collins Publishers, London.
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr. Miami 5 : 665-847.
Randall, J.E. 1985. Guide to Hawaiian reef fishes. Harrowood Books, Newtown Square, Pennsylvania. 74 p.
Scott, W.E. et E.J. Crossman. 1973. Freshwater fishes of Canada. Bull. Fish. Res. Board Can. 184, 966 p.
Sierra, L.M., R. Claro et O.A. Popova. 1994. Alimentacion y relaciones tróficas, p. 263-284. In R. Claro (éd.) Ecología de los peces marinos de Cuba. Instituto de Oceanología Academia de Ciencias de Cuba by Centro de Investigaciones de Quintana Roo, Mexico.
Whitehead, P.J.P. 1985. FAO species catalogue. Vol. 7. Clupeoid fishes of the world. An annotated and illustrated catalogue of the herrings, sardines, pilchards, sprats, anchovies, and wolf herrings. Part I. Chirocentridae, Clupeidae, and Pristigasteridae. FAO Fish. Synop. 7(125) Pt. 1 : 303 p.
Maria Lourdes D. Palomares, Pascualita Sa-a et Daniel Pauly
La connaissance du régime alimentaire d'une espèce de poisson dans une localité définie est utile pour l'évaluation de sa position et de son impact écologiques, pour la modélisation des écosystèmes (voir Encadré 18), et pour faciliter la définition des exigences alimentaires des espèces susceptibles d'être l'objet d'une aquaculture. Dans FishBase, les données de la table DIET sont aussi utilisées pour l'estimation des valeurs d'un niveau trophique des espèces (voir Encadré 21).
D'autre part, la plupart des poissons démersaux ont développé des méthodes spécialisées pour collecter leur nourriture. Les ‹ brouteurs de substrat ›, les ‹ suceurs de films nutritifs › ou ‹ les brouteurs de plantes aquatiques › vivent souvent près du fond et ont des morphologies spécialisées adaptées à leur comportement dans cet habitat (voir de Groot 1981 pour les poissons plats par exemple). Des techniques d'alimentation encore plus spécialisées sont utilisées par les poissons qui dépendent d'autres organismes pour se nourrir, comme les parasites, commensaux, nettoyeurs et nécrophages. Certaines espèces présentent des types d'alimentation ‹ variables › (voir par exemple Tiews et al. 1972 sur les habitudes alimentaires des Leiognathidae). Le choix ‹ autres › indique des habitudes très spécialisées, qui ne sont pas dans la liste prédéfinie, mais qui sont précisées dans Remarques (Remarks).
De nombreuses références de la littérature fournissent des informations sur les fréquences de présence des aliments dans les contenus stomacaux des poissons, fréquences que quelques lecteurs peuvent juger utiles de proposer sur les régimes alimentaires. Cependant, à l'exception des larves de poisson, dont les aliments sont tous uniformément petits, la fréquence de présence d'un aliment n'est pas un bon indicateur de sa contribution quantitative au régime alimentaire d'une population donnée. Par exemple, un petit copépode qui est présent dans 50 % des estomacs examinés peut contribuer beaucoup moins au régime alimentaire que les grands polychètes qui sont trouvés dans 20 % des estomacs seulement. Le grand nombre d'indices appliqués aux fréquences ne pallient pas ce défaut essentiel et embrouille plutôt le sujet. Les éditeurs et les arbitres devraient rejeter les manuscrits soumis qui traitent des contenus stomacaux sans fournir de données en poids, en volume ou en énergie.
Sources
Nous avons limité nos enregistrements aux données quantitatives qui ne souffrent pas du défaut décrit ci-dessus. De plus, ils ne sont documentés que par des études et des analyses de contenus stomacaux de spécimens capturés en milieu naturel, et pas élevés en conditions expérimentales. Ainsi, la plupart des informations dans la table DIET sont extraites de relativement peu de références, dont Stevens (1966), Randall (1967), Hobson (1974), Armstrong (1982), Sano (1984), Randall (1985), Gonzalez et Soto (1988), Laroche (1982), Sierra et al. (1994) et Valtysson (1995).
Des données sur le régime alimentaire ont été compilées pour plus de 800 espèces de poissons. Nous aimerions avoir ces données pour autant d'espèces que possible, et apprécierions de recevoir des tirés-à-part pour les espèces qui nous font défaut.
Statut
Les noms et la classification taxinomique des aliments pour plus de 1 700 enregistrements ont été vérifiés dans Taxonomic Code du National Oceanographic Data Center (NODC) (Hardy (1993), Taxonomic Authority List of the Aquatic Sciences and Fisheries Information System (de Luca 1988 ; et Barnes 1980). Des incohérences peuvent survenir dans la classification fonctionnelle de quelques aliments d'origine animale. Nous avons essayé de les réduire autant que possible, bien que sans y parvenir totalement sans doute, en inférant le groupe fonctionnel d'un aliment à partir du comportement et de l'habitat de l'espèce consommatrice.
Les champs
La table DIET comprend les champs suivants qui indiquent :
Phase : La phase du cycle de vie des spécimens de poissons étudiés parmi les choix suivants : larves ; recrues/juv. ; juv. /adultes (option par défaut quand inconnu) ; adultes.
Nombre : Le nombre de spécimens étudiés ; le pourcentage d'estomacs vides est mentionné quand il est disponible.
Localité : Le site spécifique de l'étude, et le Pays.
Mois de l'étude : Ils définissent la période de l'année pendant laquelle les spécimens ont été capturés, sous forme de champs sélectionnés. Cette information peut servir à interpréter la présence ou l'abondance de tel ou tel aliment dans l'habitat.
Remarks : Précise les informations pour les choix ‹ autres › éventuellement sélectionnés, ou pour toute autre explication ou description d'un aliment jugées utiles.
Aliment I, II, III : Pour ordonner tout l'éventail d'informations trouvées dans la littérature, les aliments sont classés en trois catégories, de groupes très généraux dans Aliment I (Food I) aux groupes taxinomiques dans Aliment III (Food III) (voir ‹ La table FOOD ITEMS › et Encadré 22, ce volume, pour les détails sur cette hiérarchie). De plus, les noms scientifiques de l'espèce constituant l'aliment et/ou d'autres informations peuvent être affichées dans un champ textuel en cliquant sur les boutons Plus (More), quand ils sont sélectionnés. Si les volumes ont été recalculés, ce champ indique aussi le pourcentage original de contribution de l'aliment au régime alimentaire.
Phase proie : Pour la nourriture animale :
phase du cycle de vie d'une proie parmi les choix suivants :
œufs ; larves ou pupes ; recrues/juv. ; juv.
/adultes ; adultes.
Pour la nourriture végétale : partie de la plante parmi
les choix suivants : racines ; tiges ;
feuilles/lames ; fruits/graines ; n.d./autres. Le
dernier choix est prévu pour les cas où la phase n'est pas
indiquée dans la source et qu'elle ne peut pas être inférée,
ou quand plusieurs phases sont consommées.
% diet : Indique le pourcentage en poids ou en volume de l'aliment dans les contenus stomacaux ; la somme des pourcentages de tous les aliments est indiquée dans un champ calculé et doit être égale à 100%. Les articles non identifiés dans le bol alimentaire sont exclus avant le calcul du pourcentage. La distribution des pourcentages peut aussi être affichée sous forme de camembert en cliquant sur l'icône ‹ graphique › dans la partie supérieure de la vue (voir Fig. 31).
Les valeurs moyennes et les erreurs-standard de troph sont indiquées dans un champ calculé à partir du régime alimentaire (et de trophs des éléments de nourriture) ; voir ‹ La table FOOD ITEMS ›).

Fig. 31. Le régime alimentaire, en % du volume ou du poids chez Oreochromis niloticus niloticus au Lac Awasa. La décomposition détaillée des catégories présentées est disponible dans FishBase.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, le bouton Trophic ecology dans la vue BIOLOGY et le bouton Diet composition dans la fenêtre TROPHIC ECOLOGY. Double-cliquer sur une des lignes dans la vue DIET COMPOSITION affiche les informations relatives à l'étude de régime alimentaire désignée.
Références
Armstrong, M.J. 1982. The predator-prey relationships of Irish Sea poor-cod (Trisopterus minutus L.), pouting (Trisopterus luscus L.), and cod (Gadus morhua L.). J. Cons. CIEM 40 : 135-152.
Barnes, R.D. 1980. Invertebrate zoology. 4ème édition. JMC Press, Inc., Quezon City, Philippines. 1089 p.
de Groot, S.J. 1984. Dutch observations on rare fish and Crustacea in 1981. Annales Biologiques (Copenhagen) 38 : 206.
de Luca, F. 1988. Taxonomic authority list. Aquatic Sciences and Fisheries Information System Ref. Ser. No. 8, 465 p.
Gonzalez, G.D. et L.A. Soto. 1988. Hábitos alimentícios de peces de depredadores del sistema lagunar Huizache-Caimanero, Sinaloa, México. Inst. Cienc. Del Mar y Limnol. Univ. Nal. Autón. México 15(1) : 97-124.
Hardy, J.D. 1993. NODC taxonomic code links biology and computerized data processing. Earth System Monitor 4(2) : 1-2.
Hobson, E.S. 1974. Feeding relationships of teleostean fishes on coral reefs in Kona, Hawaii. Fish. Bull. 72(4) : 915-1031.
Laroche, J.L. 1982. Trophic patterns among larvae of five species of sculpins (Family : Cottidae) in a Maine estuary. Fish. Bull. 80(4) : 827-840.
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr. Miami 5 : 665-847.
Randall, J.E. 1985. Guide to Hawaiian reef fishes. Harrowood Books, Newton Square, Pensylvania.
Sano, M., M. Shimizu et Y. Nose. 1984. Food habits of teleostean reef fishes in Okinawa Island, southern Japan. University of Tokyo Press, Tokyo, Japan. 128 p.
Sierra, L.M., R. Claro et O.A. Popova. 1994. Alimentacion y relaciones tróficas, p. 263-284. In R. Claro (éd.) Ecología de los peces marinos de Cuba. Instituto de Oceanología Academia de Ciencias de Cuba by Centro de Investigaciones de Quintana Roo, Mexico.
Stevens, D.E. 1966. Food habits of striped bass, Roccus saxatilis in the Sacramento-San Joaquin Delta, p. 68-96. In J.L. Turner et D.W. Kelly (compilateurs). Ecological studies of the Sacramento-San Joaquin Delta. Part II. Fishes of the Delta. Fish. Bull. 136.
Tiews, K., S.A. Bravo, I.A. Ronquillo et J. Marques. 1972. On the food and feeding habits of eight species of Leoignathus found in Manila Bay and San Miguel Bay. Indo-Pac. Fish. Counc. 13(3) : 93-99.
Valtysson, H.T. 1995. Feeding habits and distribution of eelpout species Lycodes spp. (Reinhardt) (Pisces : Zoarcidae) in Icelandic waters. Postgraduate thesis, Department of Biology, University of Iceland, Reykjavik.
Maria Lourdes D. Palomares et Pascualita Sa-a
Comme les autres organismes hétérotrophes, les poissons ont besoin de se nourrir pour survivre et croître. Dans les écosystèmes, les relations trophiques et les flux d'énergie définissent la position des diverses espèces pour une grande part (voir l'Encadré 19, et Christensen et Pauly 1993). Il y a deux façons de présenter une consommation espèce-spécifique :
Pauly (1986) et Palomares et Pauly (1989 ; 1998) discutent de la relation entre ces deux mesures et des méthodes pour l'estimation de ces paramètres. La table RATION et la table POPQB (ci-après) présentent 471 enregistrements de Rd pour 65 espèces et 161 enregistrements de Q/B pour 97 espèces, pour la plupart extraits de Palomares (1987), Palomares et Pauly (1989 ; 1998), et Pauly (1989) et Palomares (1991).
Sources
Les enregistrements proviennent pour la plupart des travaux propres ou en collaboration du premier auteur. Les sources (Référence) et les répartitions (Localité, Pays et Salinité) ont été vérifiées visuellement. Les noms et la classification taxinomique des aliments ont été vérifiés dans le Taxonomic Code du National Oceanographic Data Center (NODC) (Hardy 1993).
Les champs
Nous rappelons que le terme ‹ ration › (Rd) se rapporte à une valeur de consommation de nourriture journalière par un poisson d'une taille spécifique. Cette table présente des valeurs de ration et des paramètres afférents dans les champs suivants :
La ration journalière, le taux d'évacuation et K1 varient avec le poids du poisson étudié (Fig. 32), avec le type de nourriture ingérée, et la température moyenne (en ºC) de l'eau où vit le poisson. Les deux champs Poids du poisson (Weight of fish) et Temp. Eau (Water temp.) sont des champs numériques. Le champ Salinité (Salinity) se rapporte à 1a masse d'eau où le spécimen a été échantillonné ou étudié parmi les choix suivants : eau de mer ; eau saumâtre ; eau douce.
Le type de nourriture est décrit en utilisant deux champs à choix multiples Aliment I (Food I) et Aliment II (Food II). Aliment I (Food I) propose 6 groupes fonctionnels : détritus ; plantes ; zoobenthos ; zooplancton ; necton ; autres. Aliment II (Food II) propose des groupements plus détaillés d'aliments qui suivent la hiérarchie décrite dans la table FOOD ITEMS et l'Encadré 22. Ces deux champs incluent le choix ‹ autres › pour des articles non listés. Nom Aliment (Food name) permet de détailler la description de l'aliment, par exemple, son nom scientifique ou commun, ou sa préparation pour tous les types de nourriture artificielle telles que les boulettes (humides) ou les granulés (secs).
Les méthodes utilisées pour l'estimation du taux d'évacuation et de la ration journalière sont indiquées respectivement dans taux évac. (Evac. rate) et Ration (Ration). Le taux d'évacuation est estimé en utilisant l'une ou l'autre de deux approches générales :
Le logiciel développé à l'ICLARM pour rendre effectif le modèle de Sainsbury (1986), MAXIMS (voir Jarre et al. 1991), est maintenant largement utilisé pour la deuxième approche. Il est donc inclus comme un des choix pour le calcul du taux d'évacuation. Les autres choix possibles sont les ‹ expériences en laboratoire › comme dans (1) ci-dessus et ‹ autre › (‹ laboratory experiment › et ‹ other ›).
La méthode de calcul de la ration journalière est indiquée parmi les choix suivants : utilisation de données de contenus stomacaux avec le logiciel MAXIMS ; par le produit de taux d'évacuation et du contenu stomacal moyen (Elliott et Persson 1978) ; autres méthodes fondées sur les analyses des contenus intestinaux (par exemple Bajkov 1935 ; Gorelova 1984) ; évaluations indirectes avec le modèle métabolique de Winberg (Winberg 1956 ; Mann 1978) ; études de consommation d'oxygène (Wakeman et al 1979) ; et des expériences de nourrissage et/ou des estimations de K1 (voir Pauly 1986). Le choix ‹ autre › indique une méthode non listée. Elle doit alors être précisée dans Commentaires (Comments).

Fig. 32. Ration relative de Gadus morhua (points noirs) comparée à celle d'autres poissons. La grande dispersion est due aux différents types de nourriture, à la température environnementale et aux autres variables qui seront standardisées dans les versions futures de ce graphique.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Trophic ecology dans la vue BIOLOGY et sur le bouton Ration dans la fenêtre TROPHIC ECOLOGY. Double-cliquer sur une des lignes dans la vue LIST OF RATION STUDIES affiche les informations relatives à l'étude désignée.
Nous prévoyons que le nombre d'espèces et de stocks dans cette table augmentera dans le futur, car des ensembles de données adéquats sont disponibles, présentés notamment aux Réunions Scientifiques annuelles du Conseil International pour l'Exploration de la Mer.
Références
Bajkov, A.D. 1935. How to estimate the daily food consumption of fish under natural conditions. Trans. Am. Fish. Soc. 65 : 288-289.
Christensen, V. et D. Pauly, Éditeurs. 1993. Trophic models of aquatic ecosystems. ICLARM Conf. Proc. 26, 390 p.
Elliott, J.M. et L. Persson. 1978. The estimation of daily rates of food consumption for fish. J. Anim. Ecol. 47 : 977-993.
Gorelova, T.A. 1984. A quantitative assessment of consumption of zooplankton by epipelagic lantern fishes (Family Myctophidae) in the equatorial Pacific Ocean. J. Ichthyol. 23(3) : 106-113.
Hardy, J.D. 1993. NODC taxonomic code links biology and computerized data processing. Earth Systems Monitor 21(2) : 1-2.
Jarre, A., M.L. Palomares, M.L. Soriano, V.C. Sambilay, Jr. et D. Pauly. 1991. Some new analytical and comparative methods for estimating the food consumption of fish. ICES Mar. Sci. Symp. 193 : 99-108.
Mann, K.H. 1978. Estimating the food consumption of fish in nature, p. 250-273. In S.D. Gerking (éd.) Ecology of freshwater fish production. Blackwell Scientific Publications, Oxford.
Palomares, M.L.D. 1987. Comparative studies on the food consumption of marine fishes with emphasis on species occurring in the Philippines. Institute of Biology, College of Science, University of the Philippines, Diliman, Quezon City. 107 p. MS thesis.
Palomares, M.L.D. 1991. La consommation de nourriture chez les poissons : étude comparative, mise au point d'un modèle prédictif et application à l'étude des réseaux trophiques. Ecole Nationale Supérieure, Institut National Polytechnique de Toulouse. 211 p. Thèse de Doctorat.
Palomares, M.L. et D. Pauly. 1989. A multiple regression model for predicting the food consumption of marine fish populations. Aust. J. Mar. Freshwat. Res. 40 : 259-273.
Palomares, M.L. et D. Pauly. 1998. Predicting food consumption of fish populations as functions of mortality, food type, morphometrics, temperature and salinity. Mar. Freshw. Res. 49 : 447-453.
Pauly, D. 1986. A simple method for estimating the food consumption of fish populations from growth data and food conversion experiments. Fish. Bull. 84(4) : 827-839.
Pauly, D. 1989. Food consumption by tropical and temperate marine fishes : some generalizations. J. Fish Biol. 35 (Supplement A) : 11-20.
Sainsbury, K.J. 1986. Estimation of food consumption from field observations of fish feeding cycles. J. Fish Biol. 29 : 23-36.
Wakeman, J.M., C.R. Arnold, D.E. Wohlschlag et S.C. Rabalais. 1979. Oxygen consumption, energy expenditure and growth of the red snapper (Lutjanus campechanus). Trans. Am. Fish. Soc. 108 : 288-292.
Winberg, G.G. 1956. Rate of metabolism and food requirements of fishes. Fish. Res. B. Can. Trans. Ser. No. 194.
Maria Lourdes D. Palomares et Daniel Pauly
Pauly (1986) a introduit une approche d'estimation de la consommation de nourriture des populations, qui prend en compte la structure d'âge des populations, définie par :
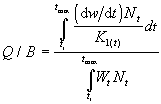 ...1)
...1)
où
![]() = 1 -
= 1 - ![]() ...2)
...2)
![]() =
= ![]() · exp
· exp ![]() ;
et ...3)
;
et ...3)
L'équation (2) implique K1 = 0 à W![]() , moment où la nourriture est utilisée
seulement pour l'entretien (Q/B de maintien) et plus pour
sa croissance. Noter que la plupart des estimations de la taille
asymptotique publiées dans la littérature se rapportent à la
longueur L
, moment où la nourriture est utilisée
seulement pour l'entretien (Q/B de maintien) et plus pour
sa croissance. Noter que la plupart des estimations de la taille
asymptotique publiées dans la littérature se rapportent à la
longueur L![]() . Une
relation longueur-poids, traduite par la constante b
(souvent fixée à 3 en l'absence d'une gamme suffisante de
paires de données L/W) est souvent utilisée pour estimer
W
. Une
relation longueur-poids, traduite par la constante b
(souvent fixée à 3 en l'absence d'une gamme suffisante de
paires de données L/W) est souvent utilisée pour estimer
W![]() à partir de L
à partir de L![]() . (voir ‹ Dynamique des
populations ›, ce volume).
. (voir ‹ Dynamique des
populations ›, ce volume).
La mortalité totale (Z) de l'équation (3) est la somme des
mortalités naturelle (M) et par pêche (F). Dans
les populations inexploitées, où F est nulle, toute la
mortalité est due à M. La température de l'eau est un
autre paramètre affectant le métabolisme et la croissance des
poissons et donc leur consommation de nourriture (Palomares et
Pauly 1989 ; Pauly 1989 ; Palomares 1991). La Température
environnementale (de l'eau) en ![]() C est par
conséquent indiquée.
C est par
conséquent indiquée.
Comme dans la table RATION, existent les champs à choix multiples Type de nourriture (Food type) et Salinité (Salinity), et un champ textuel pour la Localité (Locality). Le Type de nourriture concerne les aliments impliqués dans l'estimation de Q/B parmi les choix suivants : détritus ; plantes ; zoobenthos ; zooplancton ; necton ; autres. Le choix ‹ autres › est utilisé pour les populations qui ont été nourries de boulettes ou de granulés ou d'autres nourritures artificielles. Remarques (Remarks) contient des détails supplémentaires.
L'habitat de la population est indiqué par le type de Salinité (Salinity), eau de mer, eau saumâtre ou eau douce, et par Localité (Localité) et Pays (Country).
Un graphique présente la courbe Q/B en fonction de W![]() (voir Fig. 33).
(voir Fig. 33).

Fig. 33. Consommation relative de nourriture des poissons tropicaux (points noirs) comparée à celle d'autres espèces.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Trophic Ecology dans la vue BIOLOGY et sur le bouton Food consumption dans la vue TROPHIC ECOLOGY. Double-cliquer sur une des lignes dans la vue FOOD CONSUMPTION affiche les informations relatives au Q/B désigné.
Le développement futur de cette table pourrait impliquer des améliorations de l'équation (2) suggérées par Temming (1994). De plus, nous pensons qu'une augmentation du nombre d'enregistrements de Q/B pourrait mettre en évidence des relations généralisées dépassant celles de Pauly (1989), Palomares (1991) ou Palomares et Pauly (1998).
Références
Palomares, M.L.D. 1991. La consommation de nourriture chez les poissons : étude comparative, mise au point d'un modèle prédictif et application à l'étude des réseaux trophiques. Ecole Nationale Supérieure, Institut National Polytechnique de Toulouse. 211 p. Thèse de Doctorat.
Palomares, M.L. et D. Pauly. 1989. A multiple regression model for predicting the food consumption of marine fish populations. Aust. J. Mar. Freshwat. Res. 40 : 259-273.
Palomares, M.L. et D. Pauly. 1998. Predicting food consumption of fish populations as functions of mortality, food type, morphometrics, temperature and salinity. Mar. Freshw. Res. 49 : 447-453.
Pauly, D. 1986. A simple method for estimating the food consumption of fish populations from growth data and food conversion experiments. Fish. Bull. 84(4) : 827-839.
Pauly, D. 1989. Food consumption by tropical and temperate marine fishes : some generalizations. J. Fish Biol. 35 (Supplement A) : 11-20.
Temming, A. 1994. Food conversion efficiency and the von Bertalanffy growth function. Part I. A modification of Pauly's model. Naga, ICLARM Q. 17(1) : 38-39.
Maria Lourdes D. Palomares et Daniel Pauly
La table PREDATORS donne une liste de prédateurs référencés pour une espèce de poisson donnée. Cette table inclut le site de l'étude, le nom et une classification du prédateur ; sa phase du cycle de vie concernée ; celle de la proie et son pourcentage en poids ou en volume de contribution au régime alimentaire du prédateur. Les informations compilées dans cette table peuvent être utiles pour les halieutes et les gestionnaires de la biodiversité, parce que les relations prédateur-proie peuvent aider à expliquer la situation de certains stocks de poissons. De plus, ces informations peuvent être utilisées pour tester des hypothèses actuelles sur les tailles relatives des proies et des prédateurs (Encadré 23 ; voir aussi Figs. 34 et 35).
Sources
La table PREDATORS contient plus de 2 000 enregistrements pour plus de 800 espèces, extraits d'une centaine de références, par exemple Hiatt et Strasburg (1960), Randall (1967), Scott et Crossman (1973), Matthews et al. (1977), Ebert et al. (1981), Uchida (1981), Collette et Nauen (1983), Meyer et Smale (1991), Hensley et Hensley (1995) et Tokranov et Maksimenkov (1995). Les noms et la classification taxinomique des espèces de prédateurs autres que poissons ont été vérifiées dans Taxonomic Code du National Oceanographic Data Center (NODC) (Hardy 1993) et Taxonomic Authority List of the Aquatic Sciences and Fisheries Information System (de Luca 1988).
Les champs
Pays/localité (Country/locality) : Indique le site où l'étude a été réalisée.
Préd I (Pred. I), Préd II (Pred. II) : Indique une classification du prédateur parmi des choix présentés dans l'Encadré 23.
Groupe préd. (Pred. Group) : Indique la famille ou un taxon supérieur du prédateur.
Nom préd. (Predator Name) : Indique le nom scientifique ou commun du prédateur.
Phase préd. (Pred. Stage) : Indique la phase du cycle de vie du prédateur parmi les choix suivants : larves ; recrues/juv. ; juv./adultes ; adultes.
Phase proie (Prey Stage) : Indique la phase du cycle de vie de la proie parmi les choix suivants : larves ; recrues/juv. ; juv./adultes ; adultes.
Pour ces deux derniers champs, juv./adultes est la phase par défaut quand elle n'était pas précisée dans la référence.
% Cont. stomacaux (% of stomach contents) : Indique le pourcentage en poids ou en volume de la contribution de la proie aux contenus stomacaux du prédateur. Si un pourcentage précis n'est pas disponible, une ‹ fréquence › de la proie dans le régime alimentaire du prédateur est indiquée à côté : rare (1-5%) ; commun (6-20%) ; très commun (21-50%) ; dominant (›50%).
Remarks : Contient une description de la proie, ou la précise dans les cas ‹ autres › de Préd. I et Préd. II, ou toute autre information jugée utile.
| Encadré 23. La hiérarchie des prédateurs.
Pour standardiser la dénomination des prédateurs dans la table PREDATORS de FishBase, une classification hiérarchique a été établie, plus ou moins analogue à celle des aliments dans Food I-III de la table FOOD ITEMS (voir Encadré 22) :
Cette hiérarchie inclut en général des animaux généralement considérés comme consommateurs de poissons y compris leurs larves. Les groupes qui se nourrissent occasionnellement de poissons tels que des tuniciers qui se nourrissent sur Vinciguerria (ou bien comme les nandous, ‹ autruches ›, sud-américaines ; voir Darwin (1845), doivent être catégorisés ‹ autres › et précisés dans groupe préd. (Pred Group). Référence Darwin, C. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle. Murray, London. Maria Lourdes D. Palomares, Pascualita Sa-a et Daniel Pauly |
| Encadré 24. Les relations prédateur-proie
chez les poissons. Relier la taille des poissons à la taille de leur proie fut la première analyse confirmant la capacité de FishBase à tester des hypothèses relativement complexes, en utilisant des données qui n'avaient pas été initialement assemblées dans ce but particulier. Les hypothèses testées ici étaient :
Dans la littérature, très peu d'études sur les habitudes alimentaire indiquent la taille des organismes ingérés (d'où le fait que les tables DIET et PREDATORS ne les incluent pas). En l'absence de données de taille spécifiques à chaque étude, la taille (= la longueur) des prédateurs et des proies a été estimée comme suit :
[Noter que cette analyse ignore les cas où les proies sont des œufs et des larves, et où les prédateurs sont des larves.] Bien qu’approximatives, ces conversions conduisent à un modèle clair (voir le Fig. 28), confirmant la première partie de l'hypothèse du point (1). Mais le ratio de taille moyen prédateur/proie, bien que proche de 4, est d'environ 3,5. Peut-être plus intéressant est le second graphique (voir Fig. 29) qui montre (en unités logarithmiques) la distribution de fréquences du ratio de taille prédateur/proie. Comme il peut être constaté, cette distribution se rapproche étroitement d'une loi log normale, ce qui généralise le modèle précédemment établi sur deux espèces seulement (Ursin 1973). Ainsi, nous pouvons confirmer que les poissons préfèrent des proies-poissons mesurant un tiers à un quart de leur longueur, et que des proies-poissons mesurant la moitié de cette longueur préférentielle ou le double sont également fréquemment ingérées. Ces deux graphiques mettent aussi en évidence les cas
extrêmes, évidemment, par exemple, les grandgousiers
(Eurypharyngidae) qui peuvent consommer des proies de
leur propre taille, ou les filtreurs et brouteurs qui
consomment des proies de plusieurs ordres de grandeur
plus petits qu'eux. |

Fig. 34. Longueur du prédateur en fonction de la longueur de la proie pour diverses espèces de poissons. Voir Encadré 24 pour une interprétation de ce graphique.

Fig. 35. Distribution de fréquences du rapport prédateur-proie pour les Gadidae et diverses espèces. Voir Encadré 24 pour une interprétation de ce graphique.
Comment y arriver
Cliquer sur le bouton Biology dans la vue SPECIES, puis sur le bouton Trophic Ecology dans la vue BIOLOGY et sur le bouton Predators dans la fenêtre TROPHIC ECOLOGY. Double-cliquer sur une des lignes dans la vue PREDATORS affiche les informations relatives au prédateur désigné. Les graphiques concernant les relations prédateur-proie sont accessibles par l'icône graphique dans la vue PREDATORS ou par la fenêtre GRAPHS.
Références
Collette, B.B. et C.E. Nauen. 1983. FAO species catalogue. Vol. 2. Scombrids of the world. An annotated and illustrated catalogue of tunas, mackerels, bonitos and related species known to date. FAO Fish. Synop. (125) : 137 p.
de Luca, F. 1988. Taxonomic authority list. Aquatic Sciences and Fisheries Information System Ref. Ser. No. 8, 465 p.
Ebert, D.A., P.D. Cowley et L.J.V. Compagno. 1991. A preliminary investigation of the feeding ecology of skates (Batoidea : Rajidae) off the west coast of southern Africa. S. Afr. J. Mar. Sci. 10 : 71-81.
Goeden, G.B. 1978. A monograph of the coral trout, Plectropomus leopardus (Lacepède). Queensland Fish. Serv. Res. Bull. 1 : 1-42.
Hardy, J.D. 1993. NODC taxonomic code links biology and computerized data processing. Earth System Monitor 4(2) : 1-2.
Hensley, V.I. et D.A. Hensley. 1995. Fishes eaten by sooty terns and brown noddies in the Dry Tortugas, Florida. Bull. Mar. Sci. 56(3) : 813-821.
Hiatt, R.W. et D.W. Strasburg. 1960. Ecological relationships of the fish fauna on coral reefs of the Marshall Islands. Ecol. Monogr. 30(1) : 65-126.
Mathews, F.D., D.M. Damkaer, L.W. Knapp et B.B. Collette. 1977. Food of western north Atlantic tunas (Thunnus) and lancetfishes (Alepisaurus). NOAA Tech. Rep. NMFS SSRF –706. 19 p.
Meyer, M. et M.J. Smale. 1991. Predation patterns of demersal teleosts from the Cape south and west coasts of South Africa. 2. Benthic and epibenthic predators. S. Afr. J. Mar. Sci. 11 : 409-442.
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr. Miami 5 : 665-847.
Randall, J.E. et V.E. Brock. 1960. Observations on the ecology of epinephelinae and lutjanid fishes of the Society Islands, with emphasis on food habits. Trans. Am. Fish. Soc. 891 : 9-16.
Scott, W.B. et E.J. Crossman. 1973. Freshwater fishes of Canada. Bull. Fish. Res. Board Can. 184 : 966 p.
Tokranov, A.M. et V.V. Maksimenkov. 1995. Feeding habits of predatory fishes in the Bol'shaya River estuary (West Kamchatka). J. Ichthyol. 35(9) : 102-112.
Uchida, R.N. 1981. Synopsis of biological data on frigate tuna, Auxis thazard and bullet tuna A. rochei. NOAA Tech. Rep. NMFS Circular 436. FAO Fish. Synop. No. 12. 463 p.
Maria Lourdes D. Palomares et Pascualita Sa-a